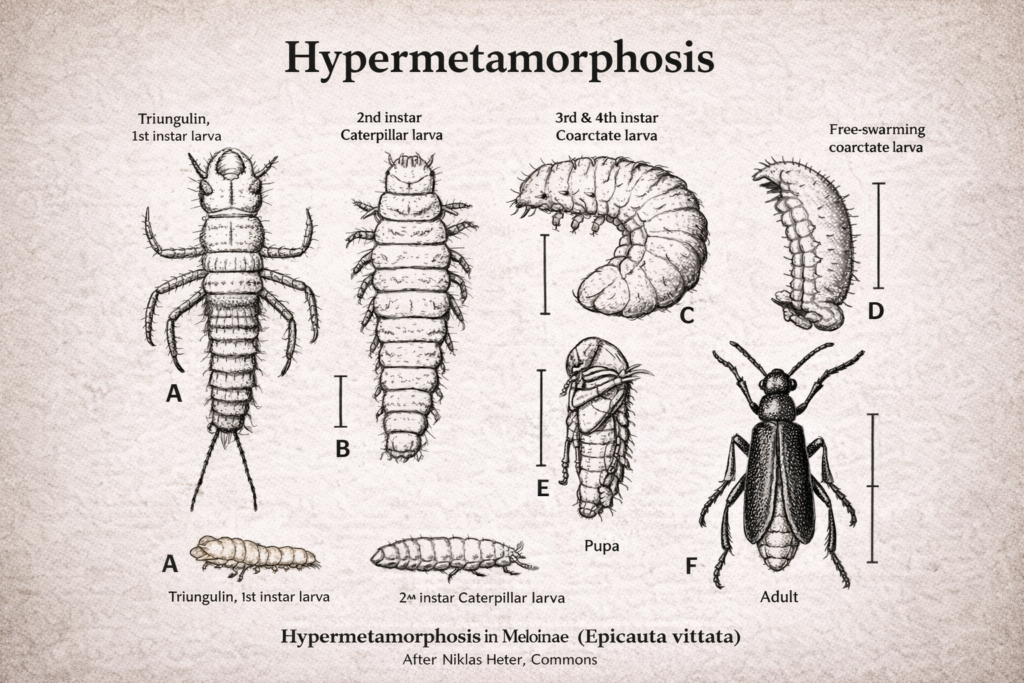
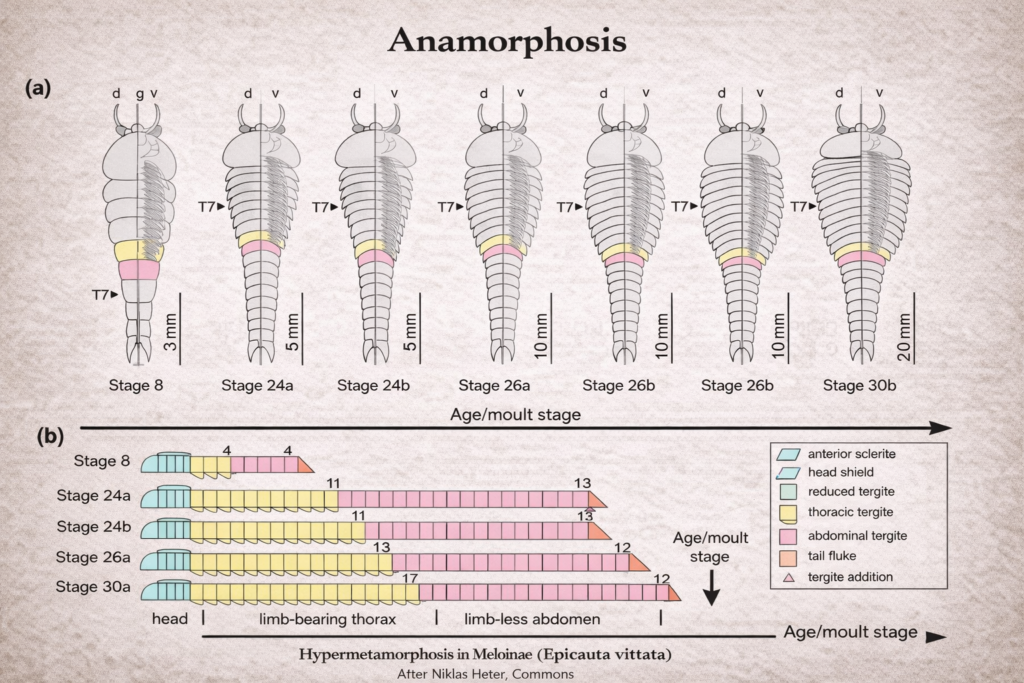
The study of insect mouthparts plays a central role in understanding the extraordinary ecological success and evolutionary diversification of insects within Class Insecta. These feeding structures are not merely passive anatomical components; instead, they function as highly coordinated, dynamic systems that allow insects to exploit an immense variety of resources. From chewing solid plant tissues to sucking nectar, blood, or plant sap, insect mouthparts demonstrate remarkable adaptability. This versatility has enabled insects to colonize nearly every terrestrial and freshwater habitat on Earth, making them the most diverse group of organisms in the animal kingdom.
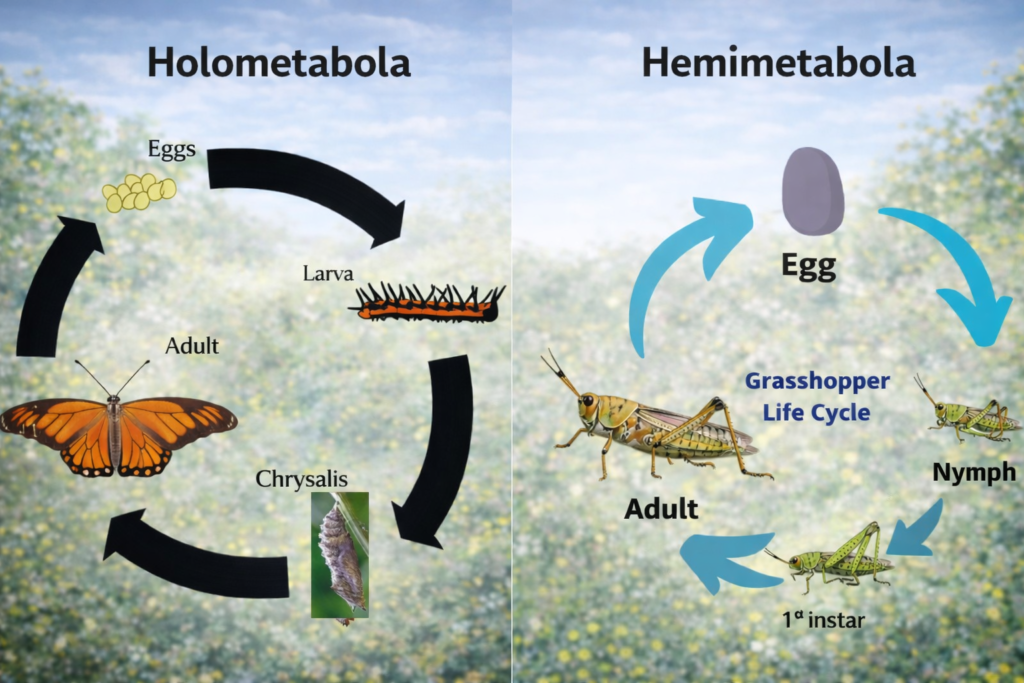
From a structural and functional perspective, insect mouthparts are classified based on their feeding mechanisms, including chewing, piercing-sucking, siphoning, and sponging types. Among these, the chewing and biting type—commonly observed in grasshoppers such as the Ak grasshopper—is considered the most primitive and generalized form. It serves as the evolutionary blueprint from which all other specialized mouthpart types have developed. By studying this basic structure, scientists can trace the morphological modifications that led to highly specialized feeding adaptations in different insect groups.
In recent years, research on insect mouthparts has advanced significantly, focusing on micromechanical efficiency, structural reinforcement through sclerotization, and integration with sensory systems. Each component, including the labrum, mandibles, maxillae, labium, and hypopharynx, is equipped with specialized grooves (sulci), joints, and sensory hairs that provide continuous feedback during feeding. This integration allows insects to precisely manipulate food, evaluate its , and optimize ingestion. Moreover, the study of insect mouthparts has practical applications in taxonomy, pest management, and agriculture, as variations in these structures can help identify species and predict their feeding behavior and economic impact.
The Five Fundamental Components of the Insect Mouth Apparatus
The structure of insect mouthparts is highly organized and consists of five primary components that work together in a coordinated manner. Each part has evolved to perform a specific function, ensuring efficient feeding and chewing processing.
Labrum (Upper Lip)
The labrum is a flap-like structure that forms the upper boundary of the mouth cavity. It is attached to the clypeus and acts as a protective covering for the internal feeding structures. However, its role is not limited to protection; it also plays an active part in feeding by holding food in position while the mandibles operate. The labrum is often flexible and capable of slight movement, allowing it to adjust according to the size and shape of the food. Additionally, it contains sensory hairs that help detect the presence and texture of food, making it an important component of insect mouthparts.
Mandibles (Primary Jaws)
Mandibles are the most and heavily sclerotized structures in insect mouthparts. These paired jaws move laterally and are responsible for cutting, crushing, and grinding food. Their allows insects to feed on твер plant materials and other substances. The mandibles are equipped with specialized зуб structures, including sharp incisor regions for cutting and broad molar regions for grinding. This dual functionality makes them highly efficient tools for mechanical digestion.
Maxillae (Accessory Jaws)
The maxillae are paired structures located beneath the mandibles and serve as accessory feeding organs. They assist in manipulating food, holding it in place, and directing it toward the mouth opening. In addition to their mechanical role, the maxillae are equipped with sensory structures that help the insect evaluate the quality and suitability of food. This sensory capability ensures that only appropriate food is ingested.
Labium (Lower Lip)
The labium forms the lower boundary of the mouth cavity and acts as a supportive platform for other mouthparts. It helps in closing the mouth cavity from the ventral side and assists in удерж food during feeding. The labium is structurally complex and includes several elements that contribute to its function. It also bears sensory appendages that enhance the insect’s ability to interact with its environment.
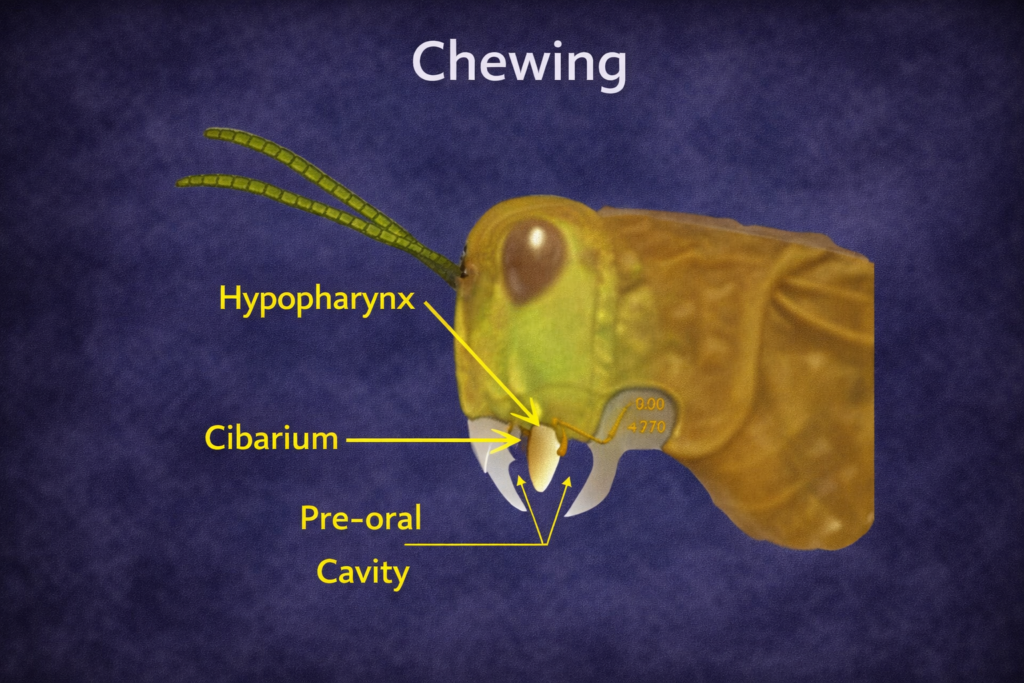
Hypopharynx (Tongue-like Structure)
The hypopharynx is a fleshy structure located enter the mouth cavity. It functions similarly to a tongue, helping to mix food with saliva and facilitating swallowing. It also plays a role in directing food toward the digestive tract. The presence of sensory hairs on the hypopharynx allows insects to detect chemical cues, further enhancing feeding efficiency.
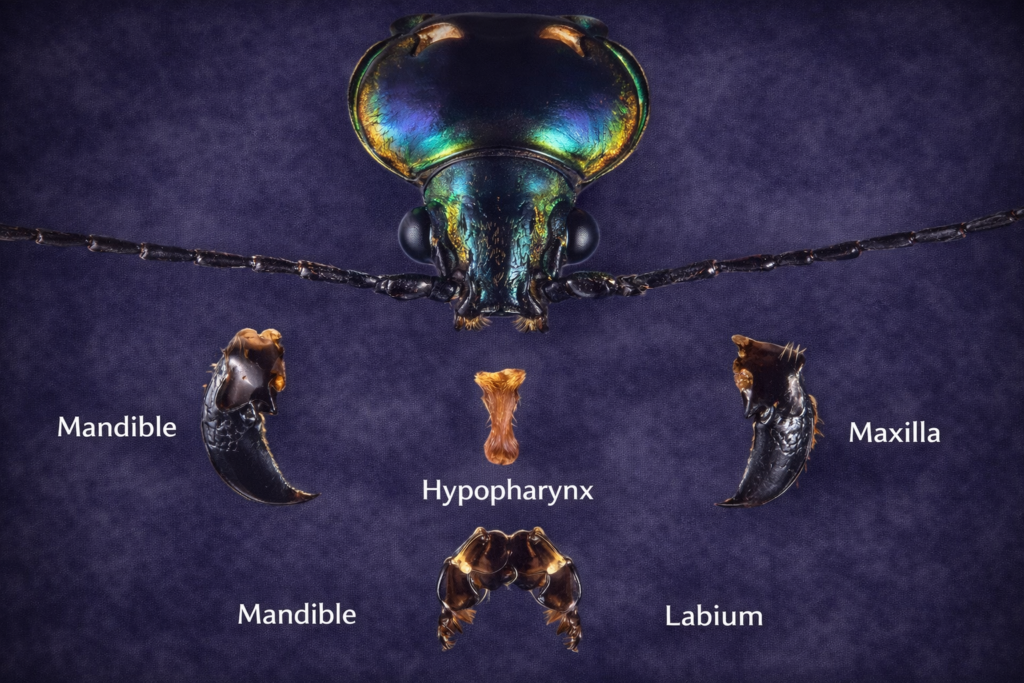
Case Study: Chewing and Biting Morphology in the Ak Grasshopper
The Ak grasshopper serves as a classical example for studying chewing-type insect mouthparts. Its feeding apparatus is well-developed and adapted for processing solid plant material, making it an ideal model for understanding the basic structure of insect feeding systems.
Dorsal vs. Ventral Views of the Labrum: Sulci and Tormae
The labrum in the Ak grasshopper is a dynamic structure capable of controlled movement.
In the dorsal view, the labrum shows two short lateral sulci and a poorly defined transverse sulcus. These grooves indicate areas of flexibility and structural division, allowing the labrum to bend and adjust during feeding.
In the ventral view, the labrum exhibits a V-shaped sulcus at the posterior region along with curved bands of sensory hairs. These sensory structures provide tactile feedback, enabling the insect to assess food texture before ingestion.
The tormae are internal, sickle-shaped supports located at the posterior angles of the labrum. They provide structural reinforcement and facilitate the movement of the labrum, ensuring smooth and coordinated feeding actions.
Mandibular Lobes: The Specialized Incisor and Molar Teeth
The mandibles of the Ak grasshopper are highly specialized and divided into two functional regions.
The incisor lobe is located at the distal end and contains sharp, pointed teeth designed for cutting and slicing plant material. This region is essential for breaking down food into manageable pieces.
The molar lobe, located at the proximal end, contains blunt teeth adapted for grinding food into smaller particles. This grinding process increases the surface area of food, making it easier for digestive enzymes to act upon it.
The presence of brustia, or hair-like structures, on the mandibles helps in удерж and guiding food particles during feeding, ensuring efficient processing.
Maxillary Sclerites: The Architecture of the Cardo and Stipes
The maxillae are structurally complex components of insect mouthparts and consist of parts that contribute to their functionality.
The cardo is the basal segment that attaches the maxilla to the head. It provides stability and allows controlled movement of the maxilla.
The stipes is attached to the cardo and serves as a platform for other maxillary structures, including the galea, lacinia, and palpi. It plays a central role in both mechanical manipulation and sensory perception of food.
Advanced Structural Analysis: Appendages and Segments
Palpi and Palpigers: The Antenna-Like Sensory Structures
Palpi are segmented, flexible appendages that function as sensory organs in insect mouthparts.
The maxillary palpus is typically five-segmented and is highly sensitive to chemical and tactile stimuli. It helps the insect evaluate food before ingestion.
The labial palpus, usually three-segmented, performs a similar function but is associated with the labium.
These structures act like miniature antennae, providing continuous sensory feedback and enhancing feeding accuracy.
Galea and Lacinia: The Apex Lobes of the Maxilla
The distal region of the maxilla bears two important lobes:
The galea is a broad and elongated structure that assists in handling and guiding food.
The lacinia is a toothed lobe that plays a role in gripping and tearing food.
When these two lobes fuse, they form a structure known as the mala, which increases functional efficiency and adaptability.
Functional Specializations: Glossae, Paraglossae, and the Ligula
The labium is divided into regions that support specialized feeding functions.
The post-mentum provides structural support and includes the submentum and mentum.
The pre-mentum contains the glossae and paraglossae, which are involved in food manipulation and handling.
The fusion of glossae and paraglossae forms the ligula, a functional structure that enhances feeding efficiency and coordination.
Sensory Integration: The Role of Sensory Hairs and Curved Bands
Sensory integration is a critical aspect of insect mouthparts, ensuring efficient and selective feeding.
The hypopharynx contains rows of sensory hairs that detect chemical properties of food, while sensory hairs on the labrum and palpi provide tactile feedback.
This combination of mechanical and sensory functions allows insects to evaluate food quality, avoid harmful substances, and optimize feeding behavior.
Conclusion
The study of insect mouthparts reveals a highly sophisticated and evolutionarily refined feeding system that has played a major role in the success of insects across diverse ecosystems. Each component, from the labrum and mandibles to the maxillae, labium, and hypopharynx, is structurally specialized and functionally integrated to ensure efficient processing. The chewing-type mouthparts, as seen in the Ak grasshopper, represent the foundational design from which more specialized feeding adaptations have evolved. Understanding these structures not only provides insights into insect biology and evolution but also has practical applications in taxonomy, pest control, and agricultural management, making it a vital area of study in modern entomology.