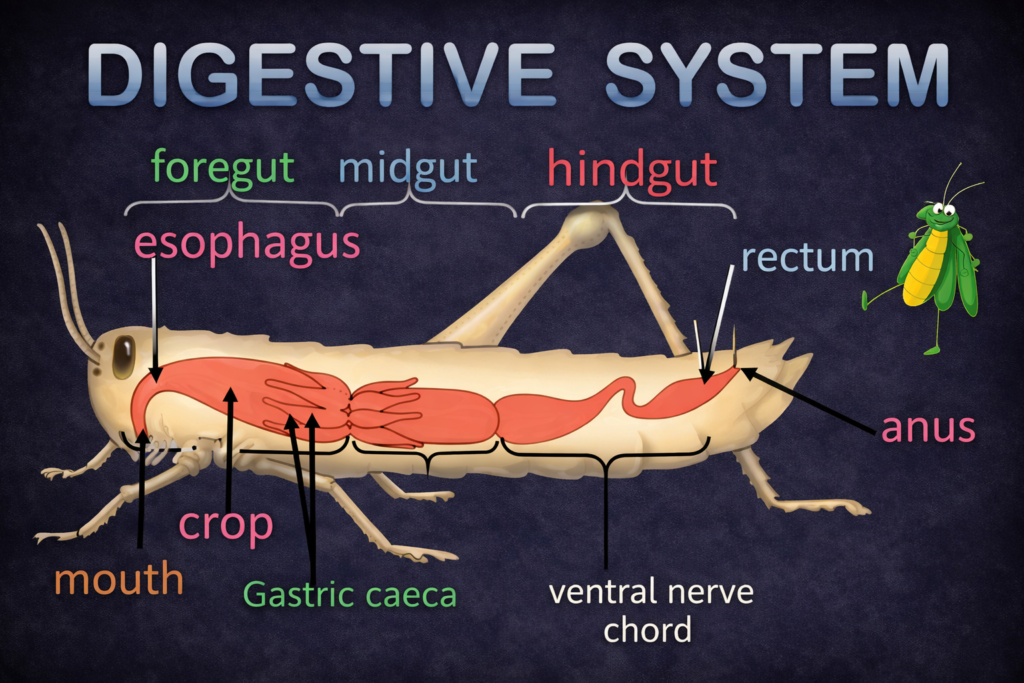
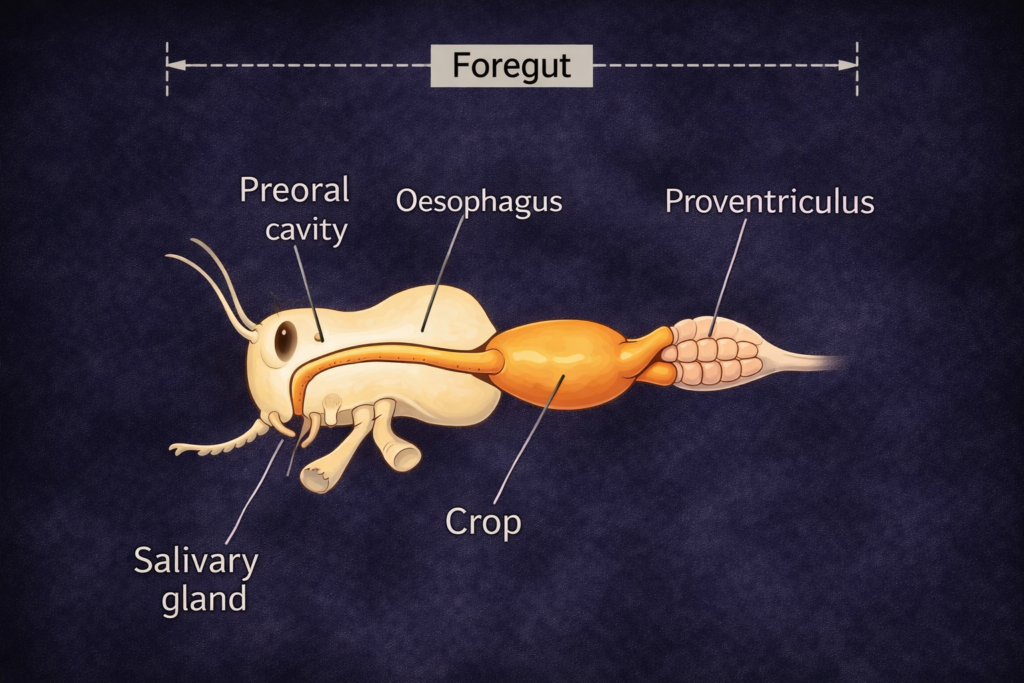
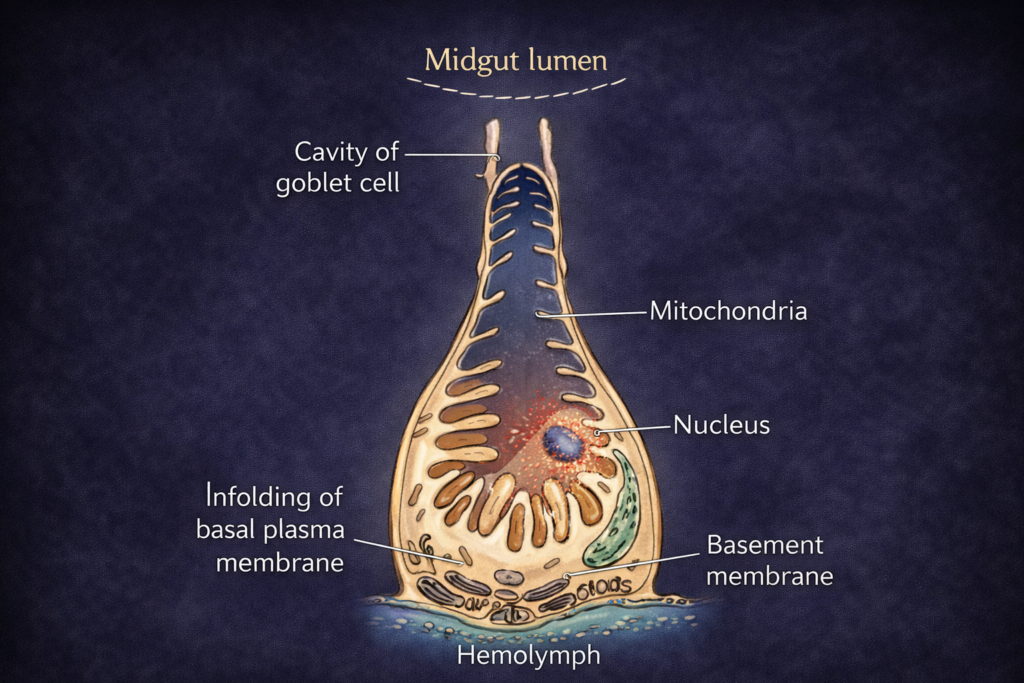
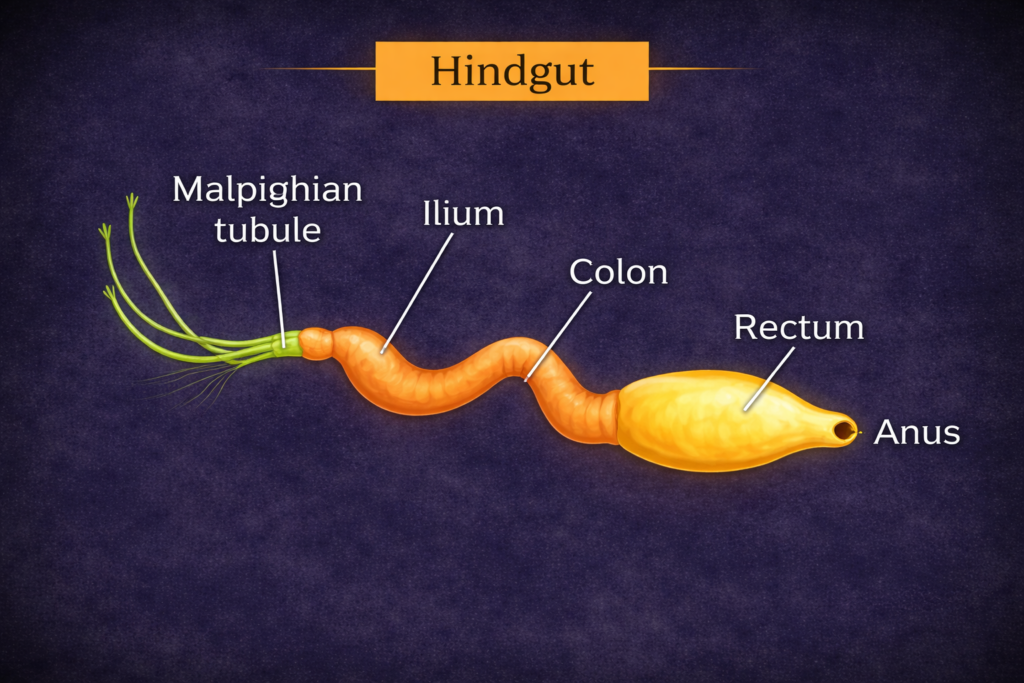
The study of Ecological Divisions is a fundamental necessity for understanding the intricate and interconnected web of life that sustains our planet. Ecology, as a scientific discipline, examines the relationships between living organisms and their physical environment. However, due to the vast scale and complexity of these interactions, a structured and systematic approach is required. By dividing ecology into well-defined Ecological Divisions, scientists are able to study life at different levels of organization, from individual organisms to entire ecosystems and the biosphere.
These Ecological Divisions provide a framework that simplifies complex biological relationships into manageable categories. Each division focuses on a specific aspect of ecological interactions, such as species behavior, population dynamics, community relationships, or environmental conditions. This structured classification allows researchers to analyze patterns, identify environmental changes, and develop predictive models. Without such divisions, it would be extremely difficult to understand how organisms survive, adapt, and interact within their environments.
In the modern era, Ecological Divisions have expanded beyond traditional boundaries due to advancements in technology and scientific methods. Tools such as remote sensing, genetic sequencing, and environmental modeling have transformed ecological research into a multidisciplinary field. Today, these divisions are not only used for academic purposes but also play a crucial role in conservation biology, climate change studies, agriculture, and natural resource management. Understanding Ecological Divisions is therefore essential for addressing global environmental challenges and ensuring sustainable development.
Core Theoretical Divisions: Autecology vs. Synecology
One of the most fundamental ways to classify Ecological Divisions is based on the number of species being studied and the scope of their interactions.
Autecology: The Study of Individual Species-Environment Dynamics
Autecology focuses on the study of a single species in relation to its environment. The term “auto” means “self,” indicating that this division deals with individual organisms or populations of the same species. It examines how a species adapts to environmental conditions such as temperature, humidity, light, and availability of food resources.
This branch of ecology is particularly important for understanding the survival strategies and life cycles of organisms. It explores how environmental factors influence growth, reproduction, and behavior. For example, studying the autecology of a pest species can help scientists determine the conditions under which it thrives and develop effective control measures.
Autecology also provides valuable insights into species-specific adaptations. These may include physiological adaptations such as tolerance to extreme temperatures, morphological adaptations like body structure, and behavioral adaptations such as migration or hibernation. By focusing on a single species, autecology allows for detailed and precise ecological analysis.
Synecology: Analyzing Aggregate Species and Community Interactions
Synecology, in contrast, focuses on the study of multiple species and their interactions within a shared environment. The term “syn” means “together,” highlighting the collective nature of this division. It examines how different species coexist, compete, and cooperate within ecological communities.
This division is essential for understanding the structure and functioning of ecosystems. It studies relationships such as predation, competition, mutualism, and symbiosis. These interactions determine the distribution and abundance of species within a community.
Synecology also investigates how communities respond to environmental changes, such as climate variation, habitat destruction, or the introduction of invasive species. By analyzing these interactions, scientists can better understand ecosystem stability and resilience. This knowledge is critical for conservation efforts and ecosystem management.

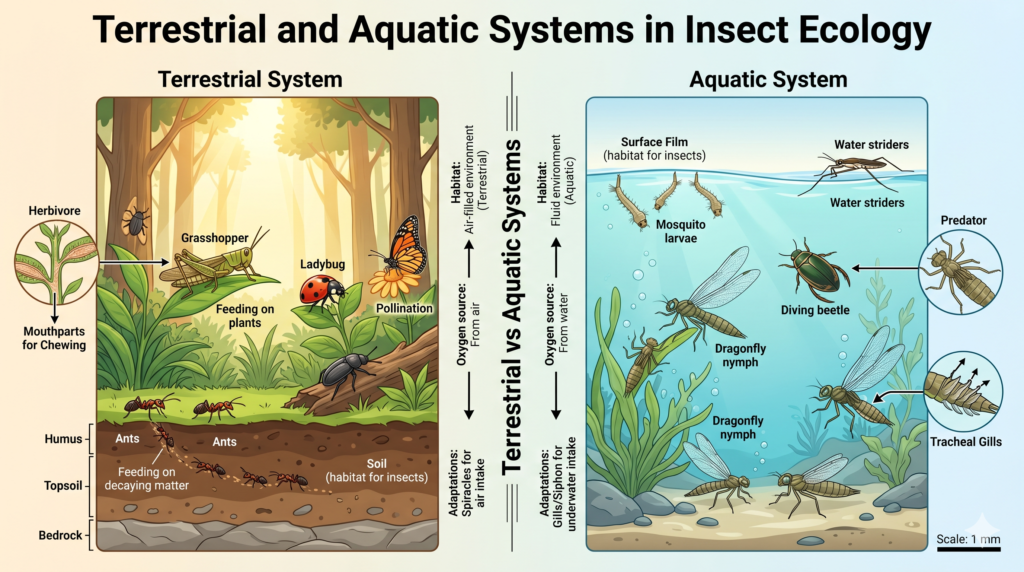
Environmental Classifications: Terrestrial and Aquatic Systems
Another important classification of Ecological Divisions is based on the physical environment in which organisms live.
Aquatic Ecology: Differentiating Freshwater and Marine Environments
Aquatic ecology focuses on ecosystems found in water bodies. These environments are divided into freshwater and marine systems, each with distinct physical and chemical characteristics.
Freshwater ecology studies ecosystems such as rivers, lakes, ponds, and wetlands. These environments have low salt concentrations and support a wide variety of organisms, including fish, amphibians, algae, and aquatic plants. Freshwater ecosystems are highly sensitive to pollution and environmental changes, making them important indicators of ecological health.
Marine ecology, on the other hand, deals with saltwater environments such as oceans and seas. These ecosystems are vast and complex, covering more than 70% of the Earth’s surface. Marine organisms have unique adaptations to survive in high salinity, pressure, and varying الضوء conditions. Marine ecology also plays a crucial role in global processes such as carbon cycling and climate regulation.
Terrestrial Ecology: Life on Land and Environmental Constraints
Terrestrial ecology focuses on life on land, where organisms face a completely different set of environmental challenges. These include variations in temperature, limited water availability, and exposure to gravity and الهواء conditions.
This division covers a wide range of ecosystems, including forests, grasslands, deserts, and tundra. Each of these environments has distinct characteristics that influence the types of organisms that can survive there. For example, desert organisms are adapted to conserve water, while forest species may be adapted to compete for light.
Terrestrial ecology also examines soil composition, nutrient cycling, and the role of vegetation in supporting life. Understanding these factors is essential for managing land resources, agriculture, and biodiversity conservation.
Organizational Hierarchy: Population and Community Levels
Ecological Divisions can also be classified based on levels of biological organization.
Population Ecology: Environmental Impacts on Cohesive Groups
Population ecology focuses on groups of individuals belonging to the same species. It studies how environmental factors affect population size, distribution, and growth.
Key aspects of population ecology include birth rates, death rates, immigration, and emigration. These factors determine whether a population grows, declines, or remains stable. Environmental resistance, such as limited resources or predation, also plays a significant role in regulating population size.
Population ecology is particularly important in agriculture and pest control. By understanding population dynamics, scientists can develop strategies to manage harmful species and protect beneficial ones.
Community Ecology: Interactions Within Multi-Species Assemblages
Community ecology examines the interactions between different species living in the same area. It focuses on how these interactions influence the structure and diversity of communities.
This division studies relationships such as competition for resources, predator-prey dynamics, and cooperative interactions. It also explores how communities change over time through processes such as ecological succession.
Community ecology is essential for understanding biodiversity and ecosystem stability. It helps scientists identify keystone species, which play a critical role in maintaining the balance of ecosystems.
Subject-Based Specializations in Ecological Study
Modern science has introduced specialized Ecological Divisions based on specific scientific disciplines.
Morphoecology and Physioecology: Structural and Functional Adaptations
Morphoecology studies how the physical structure of organisms is influenced by their environment. For example, the shape of leaves in plants or the body structure of animals can reflect adaptation to environmental conditions.
Physioecology focuses on the physiological processes that enable organisms to survive and function in their environments. This includes processes such as respiration, metabolism, and temperature regulation.
Genetic Ecology and Bioecology: The Molecular Perspective
Genetic ecology examines how environmental factors influence genetic variation and evolution. It studies how natural selection shapes the genetic makeup of populations over time.
Bioecology integrates biological and environmental studies to provide a comprehensive understanding of life processes. It combines aspects of physiology, behavior, and environmental science.
Chemical Ecology: The Role of Chemical Signaling
Chemical ecology is a specialized field that studies how organisms use chemical substances to interact with each other and their environment. These chemicals may include pheromones, toxins, and signaling compounds.
This division is particularly important in understanding insect behavior, plant defense mechanisms, and ecological communication systems.
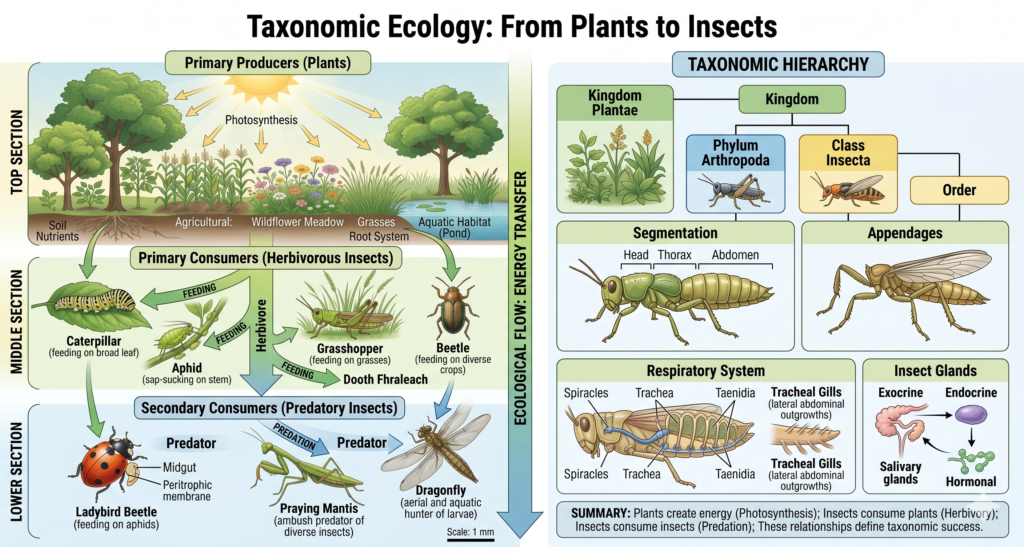
Taxonomic Ecology: From Plants to Insects
Ecological Divisions are also categorized based on the type of organisms being studied.
Plant Ecology: Understanding Vegetation Systems
Plant ecology focuses on the relationship between plants and their environment. It includes the study of different plant groups, such as gymnosperms and angiosperms, and their ecological roles.
Plants are primary producers and form the foundation of most ecosystems. Understanding plant ecology is essential for studying energy flow and nutrient cycling.
Animal Ecology: Vertebrates and Invertebrates
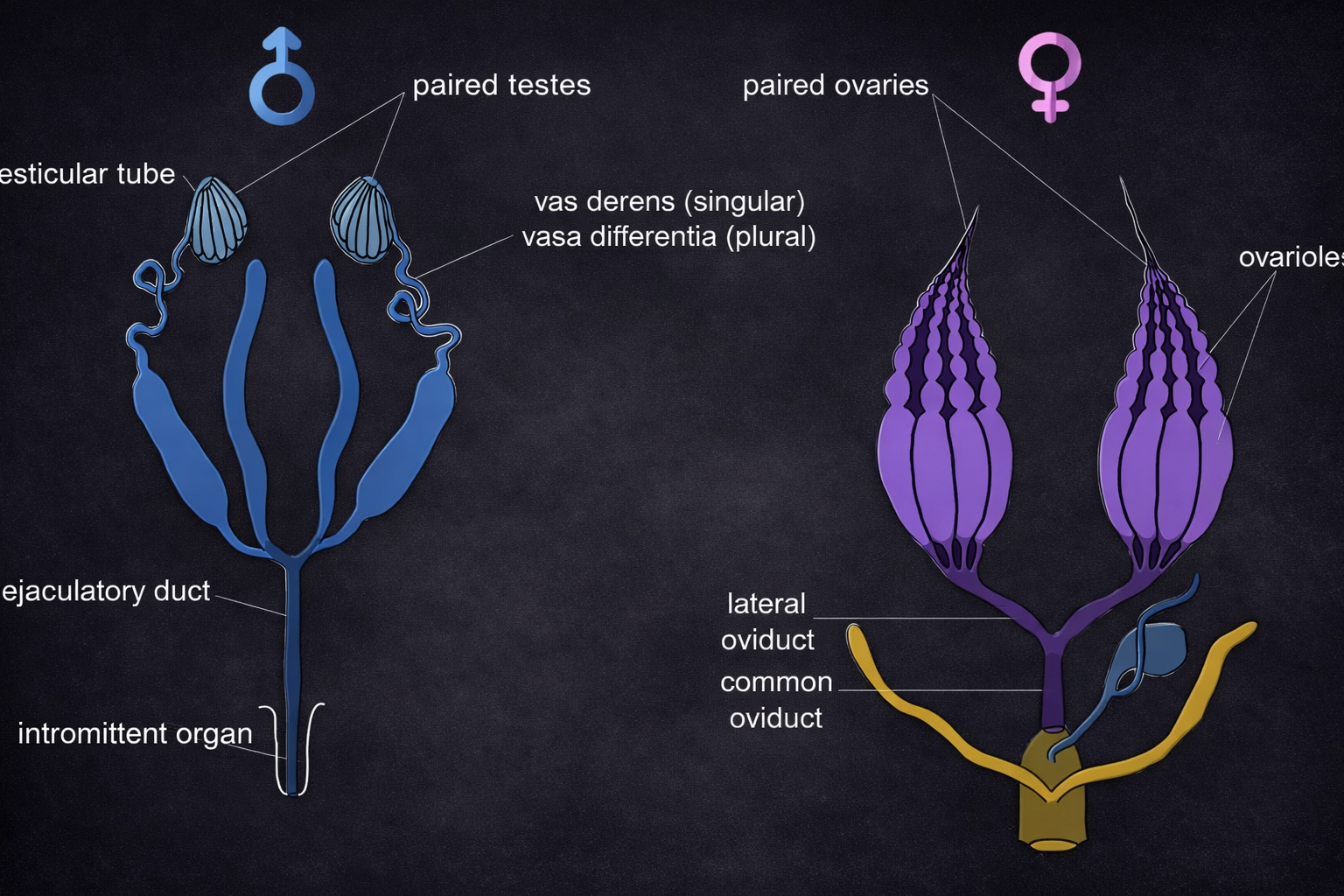
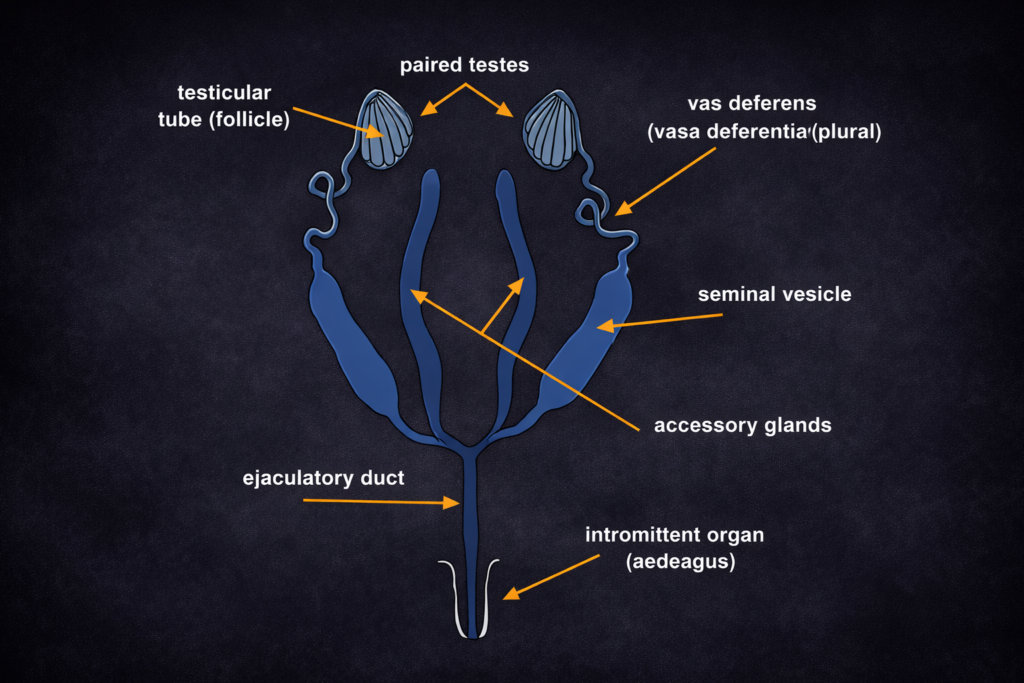
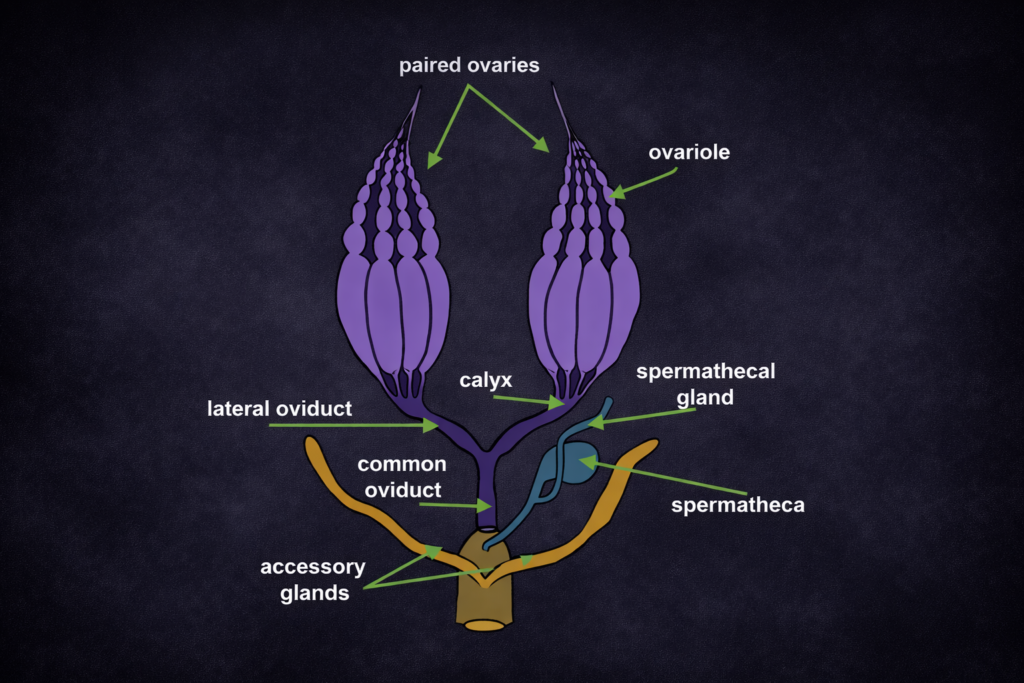
Animal ecology studies the interactions between animals and their environment. It is divided into vertebrate and invertebrate ecology.
Vertebrate ecology focuses on animals with a backbone, such as mammals, birds, and reptiles. Invertebrate ecology covers a wide range of organisms, including insects, worms, and mollusks.
Insect Ecology: A Specialized Branch
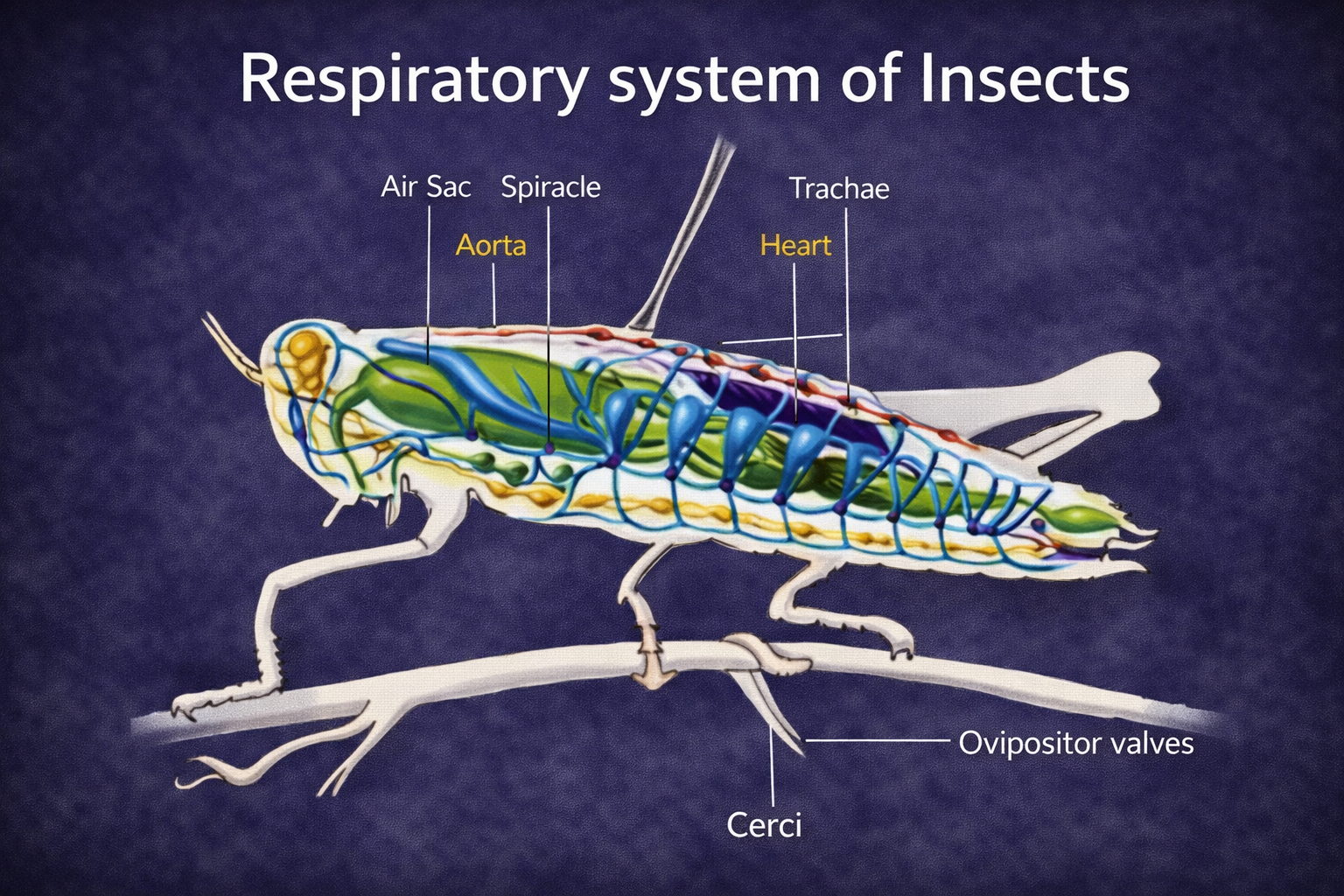
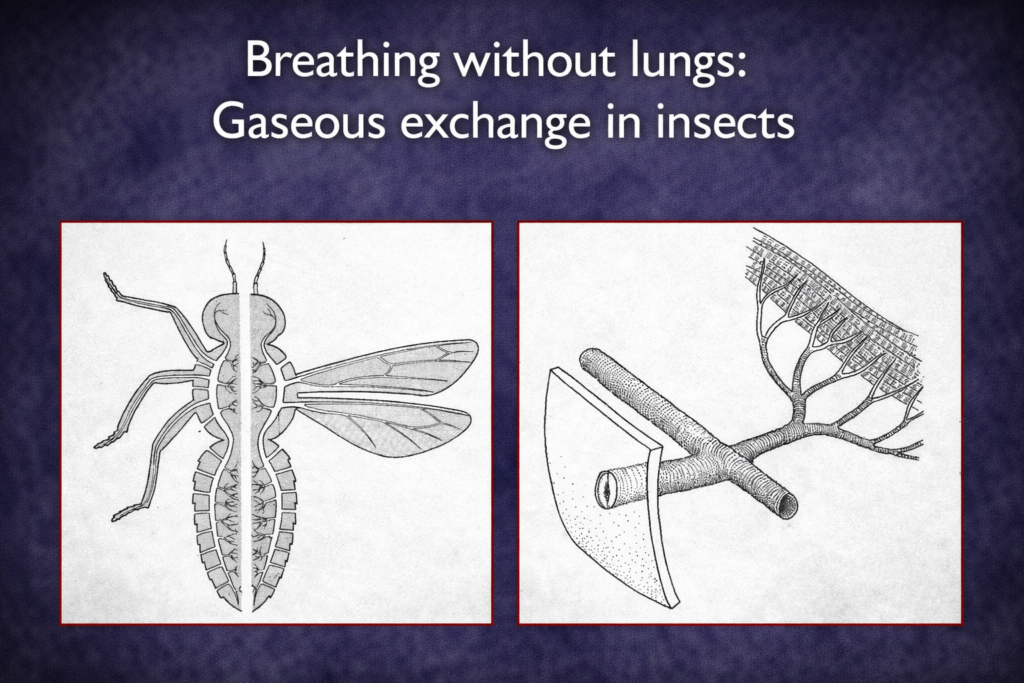
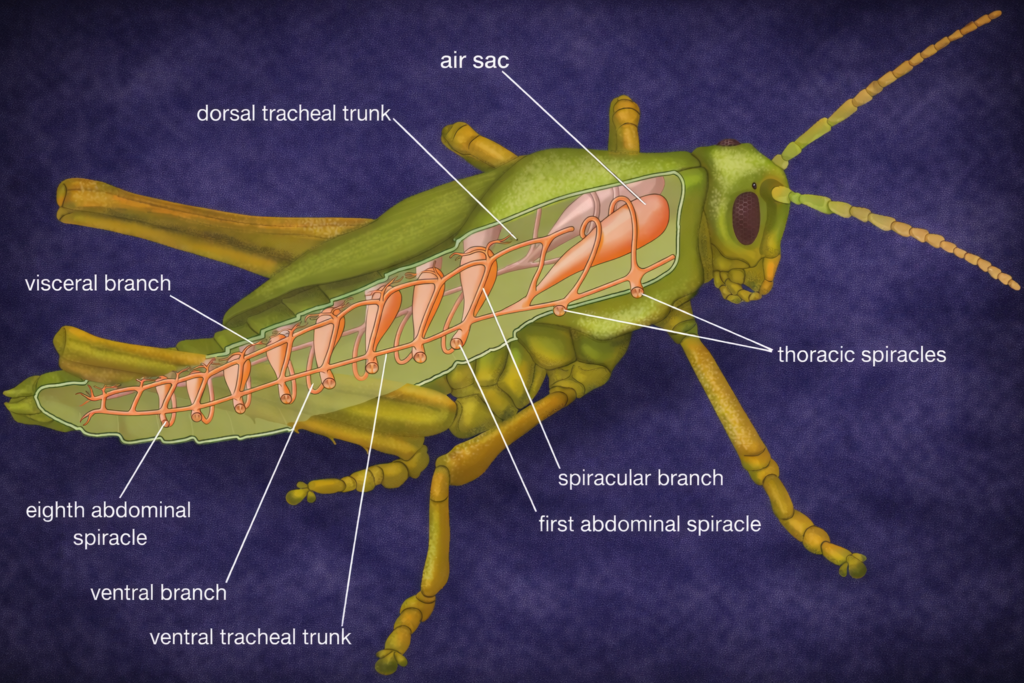
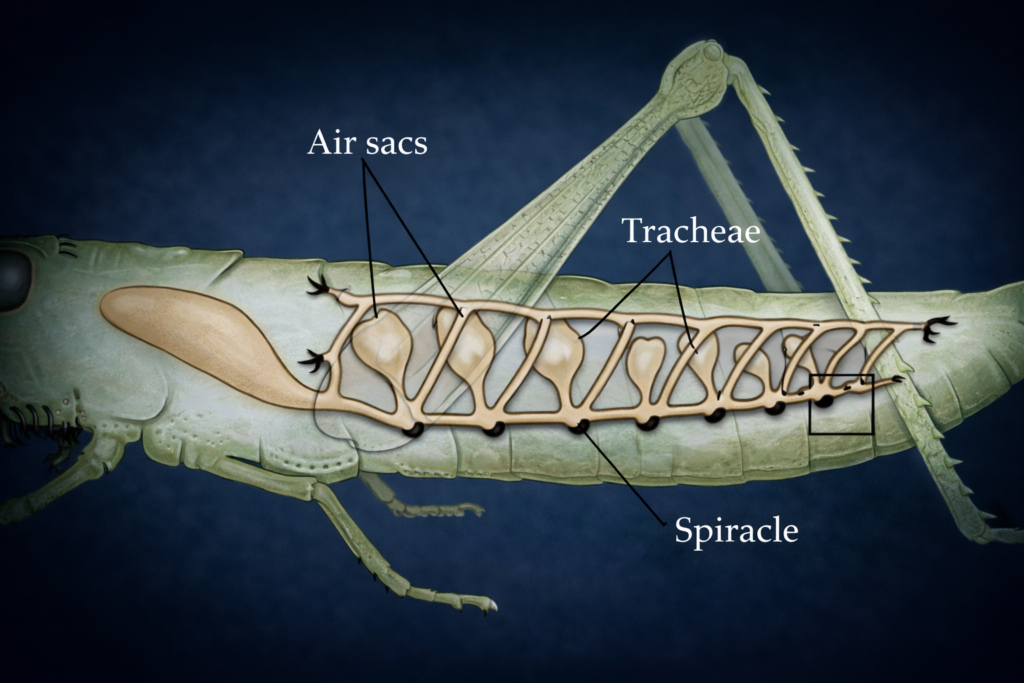
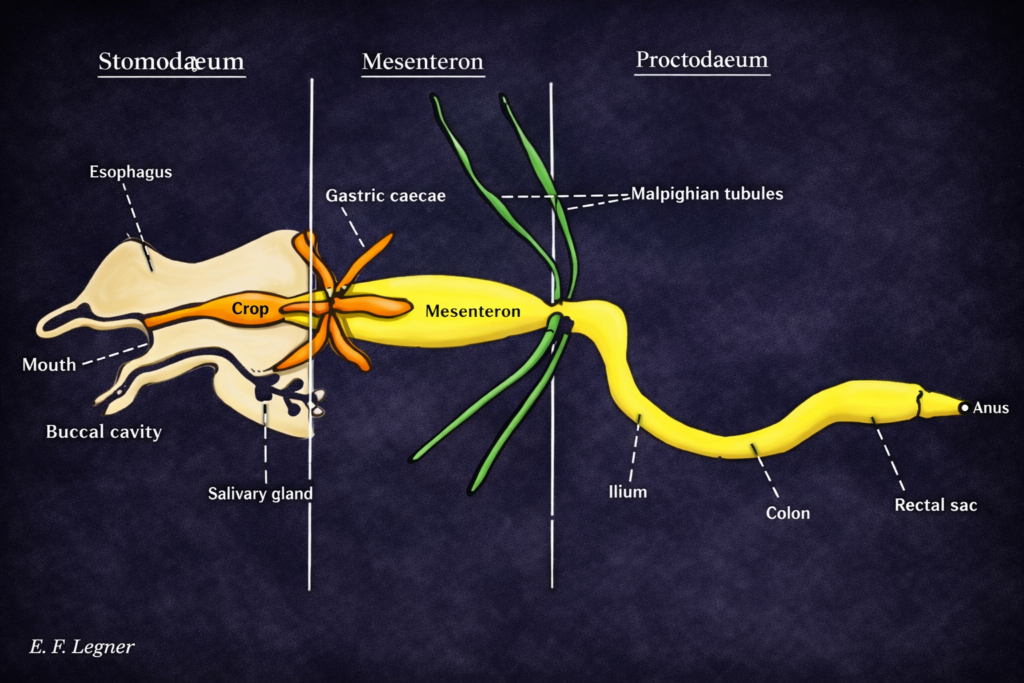
Insect ecology is a highly specialized field due to the immense diversity and ecological importance of insects. It examines how insects interact with their environment, adapt to different conditions, and influence ecosystems.
Insects play key roles as pollinators, decomposers, and pests. Understanding insect ecology is essential for agriculture, biodiversity conservation, and ecosystem management.
Conclusion
The concept of Ecological Divisions provides a structured and comprehensive framework for studying the complexity of life on Earth. By organizing ecological knowledge into different branches—ranging from individual species studies to ecosystem-level interactions—scientists can better understand how organisms survive, interact, and adapt to their environments. These divisions are essential for addressing modern environmental challenges, including climate change, habitat destruction, and biodiversity loss. Ultimately, the integration of all Ecological Divisions enables a holistic approach to conservation and sustainable resource management, ensuring the long-term stability of natural systems.
FAQs: Understanding the Branches of Ecological Science
What is the difference between Autecology and Synecology?
Autecology studies a single species, while Synecology studies multiple species and their interactions.
Why are Aquatic and Terrestrial Ecology separate divisions?
Because water and land environments have different physical conditions requiring different adaptations.
What does Population Ecology focus on?
It studies the dynamics of a group of individuals of the same species.
Is Insect Ecology part of a larger division?
Yes, it is a sub-division of invertebrate ecology within animal ecology.
What is Chemical Ecology?
It studies how chemical signals influence interactions between organisms.